ODPORNOŚĆ CHWASTÓW NA HERBICYDY
Tadeusz Praczyk
Instytut Ochrony Roślin – Państwowy Instytut Badawczy
Definicje
Odporność na herbicydy jest to dziedziczna zdolność rośliny do przeżycia i reprodukcji po jej ekspozycji na działanie herbicydu w dawce powodującej zniszczenie innych naturalnie występujących osobników danego gatunku . Cecha odporności może pojawić się w sposób naturalny, bądź może być wprowadzona za pomocą różnych technik, np. inżynierii genetycznej (Weed Science Society of America, 2020).
Innymi słowy można powiedzieć, że odporność chwastów na herbicydy jest to zdolność uprzednio wrażliwej populacji do przeżycia całego cyklu rozwojowego po zastosowaniu herbicydu w zalecanej dawce, w danych warunkach agronomicznych (Heap and LeBaron 2001).
Odporność krzyżowa jest to odporność chwastu na substancje czynne herbicydów należące do różnych grup chemicznych, ale wykazujących ten sam mechanizm działania.
Odporność wielokrotna jest to odporność chwastu na dwa lub więcej mechanizmów działania herbicydów.
Dwa główne typy odporności chwastów na herbicydy:
- Odporność w miejscu działania herbicydu (Target Site Resistance - TSR) – mutacje w specyficznych białkach
- Odporność poza miejscem działania herbicydu (Non Target Site Resistance - NTSR) – zmniejszone pobieranie i przemieszczanie substancji czynnej, zwiększony metabolizm s.c. w roślinach, sekwestracja (związanie s.c. przez inną substancję).
Określenia “odporność na herbicyd” oraz „tolerancja na herbicyd” mają całkowicie odmienne znaczenia. Otóż tolerancja na herbicyd jest dziedziczną zdolnością danego gatunku rośliny do przeżycia i reprodukcji po zastosowaniu herbicydu w zalecanej dawce. Ta właściwość nie jest wynikiem presji selekcyjnej ani manipulacji genetycznej, jest to naturalna cecha danej rośliny. Na przykład rośliny pszenicy ozimej są tolerancyjne na herbicyd 2,4-D, rośliny fiołka polnego są tolerancyjne na herbicyd fenoksaprop-P etylu.
Biotyp (gr. bios „życie”, typos „odbicie, obraz”) - osobniki różniące się od innych blisko spokrewnionych określonymi cechami biologicznymi.
W niniejszym opracowaniu określenie biotyp stosowane jest dla osobników tego samego gatunku chwastu różniących się wrażliwością na herbicydy.biotyp
Historia odporności
Jednym z istotnych problemów związanych ze stosowaniem środków ochrony roślin jest zjawisko uodporniania się szkodników, grzybów chorobotwórczych oraz chwastów na te środki. Pierwsze doniesienia na ten temat pochodzą z 1908 roku, kiedy to stwierdzono uodpornienie się niektórych owadów na stosowaną wówczas w ochronie roślin ciecz siarkowo-wapienną. Problem uodporniania się grzybów chorobotwórczych na fungicydy pojawił się w 1940 roku. Zjawisko odporności chwastów na herbicydy zaobserwowano w późniejszych czasach, co było związane z faktem, że chemiczne środki chwastobójcze zaczęto stosować na szeroką skalę dopiero w latach pięćdziesiątych XX wieku. Pierwsze udokumentowane informacje o odporności chwastów na herbicydy pochodzą z końca lat pięćdziesiątych XX wieku i dotyczą biotypów marchwii zwyczajnej (Daucus carota) odpornej na 2,4-D (Switzer 1957). Kolejne doniesienie na ten temat pochodzi z 1968 roku również z USA, gdzie stwierdzono przypadki braku skutecznego działania symazyny na rośliny starca zwyczajnego (Senecio vulgaris). Problem uodporniania się chwastów na herbicydy z biegiem czasu nasilał się. W końcu lat osiemdziesiątych XX wieku, a więc po upływie niespełna dekady od wprowadzenia na rynek herbicydów z grupy pochodnych sulfonylomocznika (inhibitory syntazy acetylomleczanowej – ALS), pojawiły się USA biotypy niektórych chwastów wykazujące odporność na te związki. W tym czasie (1985 r.) stwierdzono w Australii także pierwsze przypadki odporności chwastów jednoliściennych na herbicydy, których mechanizm działania polega na blokowaniu syntezy acetylokoenzymu A (ACCase). W latach dziewięćdziesiątych XX wieku odporność chwastów dotyczyła w głównej mierze herbicydów będących inhibitorami fotosyntezy w fotosystemie II, bo w tej grupie odporność dotyczyła aż 83 gatunków (63 biotypy odporne na triazyny i 20 odpornych na pochodne mocznika). Sytuacja zmieniła się od 2000 roku, kiedy to największym problemem stała się odporność chwastów na inhibitory ALS (grupa sulfonylomocznika).
Pierwsze doniesienie o odporności chwastów na glifosat pochodzi z końca lat 90-tych XX w. (około 20 lat po wprowadzeniu glifosatu na rynek) z Australii, gdzie stwierdzono biotypy odporne życicy sztywnej (Lolium rigidum). Pierwsze przypadki odporności na glifosat były związane z wieloletnim stosowaniem tego herbicydu w sadach lub na polach przed siewem roślin. Wprowadzenie do uprawy roślin genetycznie modyfikowanych tolerancyjnych na glifosat było uważane za doskonałe rozwiązanie narastającego problemu odporności chwastów na herbicydy z grupy inhibitorów ALS. Dzisiaj wiemy, że tak duża presja selekcyjna, gdy miliony hektarów traktowanych jest jednym herbicydem i to przez wiele lat, musiała doprowadzić do uodpornienia się chwastów. Pierwsze przypadki odporności chwastów na glifosat związane z uprawą roślin GMO odpornych na ten herbicyd stwierdzono w USA w 2001 r. W 2016 r. w świecie (na 6 kontynentach) już 35 gatunków chwastów wytworzyło biotypy odporne na ten herbicyd.
Aktualna sytuacja (2020 r.)
Obecnie w świecie ponad 260 gatunków chwastów wytworzyło biotypy odporne na herbicydy. Chwasty odporne występują w 70 krajach. Największa liczba gatunków z odpornością występuje w USA (165), w Australii (95), w Kanadzie (68), we Francji (55), w Brazylii (50), w Chinach (45) i w Hiszpanii (39). W Polsce udokumentowano obecność biotypów odpornych u 22 gatunków chwastów (czytaj więcej). Chwasty najczęściej uodporniają się na herbicydy należące do inhibitorów ALS (Grupa B wg. HRAC) – 159 gatunków, a w dalszej kolejności na herbicydy z grupy inhibitorów fotosyntezy (grupa C wg. HRAC) – 105 gatunków, z grupy inhibitorów ACCase (Grupa A wg. HRAC) – 48 gatunków, z grupy inhibitorów EPSP (glifosat) – 35 gatunków.
Mechanizmy odporności
Wiele czynników, jak na przykład sposób uprawy gleby, zmianowanie, nawożenie, nawadnianie, może powodować zmiany w populacjach chwastów. Jednak stosowanie herbicydów jest prawdopodobnie tym czynnikiem, który powoduje najszybsze zmiany w tych populacjach. Większość badaczy zajmujących się problemem odporności chwastów na herbicydy uważa, że zjawisko to nie jest efektem mutacji spowodowanych stosowaniem herbicydów, ale jest skutkiem selekcji naturalnych mutacji, które już wcześniej zaszły w populacji chwastów. Pomimo, że poszczególne rośliny w danej populacji zewnętrznie mogą nie różnić się pomiędzy sobą, to jednak na poziomie genetycznym różnice istnieją. Wieloletnie stosowanie herbicydów powoduje systematyczne eliminowanie z populacji osobników wrażliwych, co stwarza warunki do rozprzestrzeniania się biotypów odpornych i po pewnym czasie stają się one elementem dominującym w populacji. Proces selekcji biotypów odpornych przedstawiono schematycznie na rysunku 1.

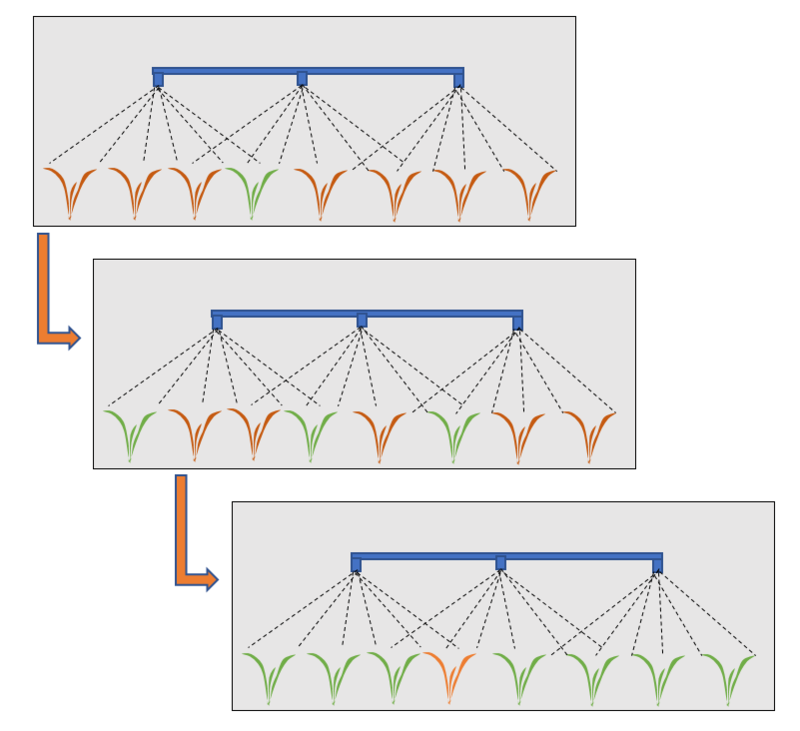
Rys. 1. Selekcja biotypów odpornych na herbicydy spowodowana stosowaniem w kolejnych sezonach herbicydów o takim samym mechanizmie działania
Mechanizmy odporności, czyli sposoby jakimi dany biotyp „unika” skutków działania herbicydu, można podzielić na dwie główne kategorie, a mianowicie:
- zmiany w miejscu działania herbicydu (odporność w miejscu działania),
- zmniejszenie możliwości przenikania herbicydu do miejsca jego działania w roślinie (odporność poza miejscem działania).
Zmiany w miejscu działania herbicydu (np. dla triazyn jest to białko D1, a dla herbicydów sulfonylomocznikowych - enzym syntaza acetomleczanowa) mogą polegać na zmianie sekwencji aminokwasów w białku, co uniemożliwia przyłączenie się substancji aktywnej do tego białka i w efekcie prowadzi do braku działania herbicydu. Zmiany takie mogą być spowodowane mutacją genu kodującego syntezę białka, z którym dany herbicyd zwykle się wiąże. Odporność na herbicydy triazynowe u większości gatunków jest wynikiem pojedynczej mutacji w genie psbA, który koduje syntezę białka D1. Efektem tej mutacji jest zamiana seryny 264 na glicynę, co powoduje utratę miejsca przyłączania się herbicydu do białka na skutek braku połączeń hydrogenowych i aminowych. Mutacje taką stwierdzono miedzy innymi u następujących roślin: Amaranthus hybridus, Solanum nigrum, Chenopodium album, Phalaris paradoxa, Brassica napus. Tego typu zmiany w genomie mogą powodować zakłócenia w procesach fizjologicznych roślin. Na przykład u biotypów odpornych na triazyny stwierdza się zmniejszenie natężenia fotosyntezy. Odporność chwastów na herbicydy z grupy inhibitorów ALS warunkowana jest mutacją, która występuje najczęściej w miejscu Pro 197, ale mogą to być także mutacje w innych miejscach (Ala 122, Ala 205, Asp 376, Arg 377, Trp 574, Ser 653, Gly 654).
Zmniejszenie możliwości przenikania herbicydu do miejsca działania może się odbywać w różnoraki sposób: ograniczenie pobierania substancji aktywnej poprzez modyfikacje niektórych cech morfologicznych (np. biotypy odporne mogą wytwarzać grubszą warstwę wosku kutikularnego lub posiadać mniejszą powierzchnię liści), ograniczenie przemieszczania się substancji czynnejj poprzez zmiany w budowie floemu i ksylemu, kompartmentacja metabolizmu, czyli rozgraniczenie przestrzenne szlaków metabolicznych (sekwestracja), czego następstwem może być zmniejszenie ilości substancji aktywnej docierającej do miejsca działania, metaboliczna detoksykacja substancji aktywnej zanim przemieści się ona do miejsca działania w roślinie (zwiększony metabolizm).
W obrębie jednej populacji chwastów mogą funkcjonować obydwa mechanizmy odporności, to znaczy zarówno odporność w miejscu działania, jak i odporność poza miejscem działania herbicydu. Na przykład zidentyfikowano, że odporności na glifosat może być spowodowana pojedynczą mutacją w genie kodującym syntazę EPSP, ale także powodem tej odporności może być sekwestracja lub redukcja przemieszczania herbicydu w roślinach.
Największym problemem jest uodpornienie się chwastów na wiele mechanizmów działania herbicydów, jak na przykład życica sztywna (Lolium rigidum) wytworzyła biotypy odporne na 14 mechanizmów działania herbicydów, chwastnica jednostronna (Echinochloa crus-galli) – na 11, a wiechlina roczna (Poa annua) na 10 mechanizmów działania (czytaj więcej). Warto w tym miejscu zaznaczyć, że wszystkie obecne aktualnie na rynku światowym herbicydy reprezentują zaledwie 25 mechanizmów działania. W tej sytuacji możliwości zwalczania chwastów z odpornością wielokrotną są bardzo ograniczone. W skali świata najwięcej gatunków chwastów odpornych na herbicydy należy do rodziny wiechlinowatych (Poaceae) , a następnie do złożonych (Asteraceae), kapustowatych (Brassicaceae), szarłatowatych (Amaranthaceae).
Różnice we wrażliwości na herbicydy pomiędzy biotypami danego gatunku chwastu określa się najczęściej przez porównanie wysokości dawki herbicydu koniecznej do zniszczenia 50% roślin (LD50), redukcji biomasy roślin (GR50 lub ED50) lub aktywności specyficznego enzymu (I50) o 50% w porównaniu do roślin nie traktowanych danym herbicydem. Bardziej precyzyjne informacje na temat wrażliwości poszczególnych biotypów można uzyskać posługując się odpowiednimi narzędziami statystycznymi w analizie zakresu dawek herbicydu, na które reagował dany biotyp.